

We will apply 3D spatial anisotropy and fractal analysis in addition to the standard Sholl analysis to characterize the branching pattern of the 3D dendritic structure of individual CA1 pyramidal cells filled with biocytin in 400 to 500 micrometer thick slices from the brains of young and aged rhesus monkeys. Individual cells will be filled with biocytin during in vitro neurophysiological recording. Then the 3D cell will be reconstructed by tracing and recording the spatial (x, y, z) positions of the soma perimeter as well as each individual dendrite by using the motorized stage and tracing algorithms of the neuron tracing module of Neurolucida.
One of the major goals in neurobiology is morphological analysis of neuronal dendritic and axonal structure. The most standard approach is applying Sholl analysis [1.] to quantify neuronal dendritic structure. The Sholl analysis of a given dendritic cell consists of determining the number of intersections of dendritic branches with spherical shell with the center at the cell body and with radius r. Sholl [1.] plotted the values either as log-log or as log-linear graph. He then used the slope of one of the two graphs to characterize the morphology of studied neurons, depending upon which of the two graphs showed linear dependence.
Some of the problems with such an analysis were pointed out in [2.]: The slope found in Sholl graphs shows big fluctuations for the cells of the same type. Thus the authors in [2.] proposed to use fractal analysis which yields fractal dimension that is related to the slope of the log-log Sholl graph. The fractal analysis consists of determining how the mass of the object scales with its size. There are several different ways to calculate the fractal dimension that can be roughly classified as length methods or mass methods, an excellent overview is given in Ref. [5.]. An advantage of fractal analysis over Sholl analysis is in the fact that the fractal dimension is not a slope of only one log-log graph, e.g. mass versus size, but it is a slope of an average log-log graph, e.g. any point on the structure is taken as a "center" from which the mass versus size is calculated. This yields less variability within the class of cells that are morphologically similar.
Fractal analysis, however, can be meaninfully applied only on those cells that have fractal structure, which may not always be the case [3.]. Nonetheless, many applications, in particular in two dimensions, of fractal analysis and have been though successfully used for morphological differentiation of different type of cells [4.,5.]. There is another problem related to morphological analysis here: Two structures may have exactly the same fractal dimension, but may be obviously different. Thus, fractal dimension by itself is not enough to completely characterize the structure of a complex cells. Another measure that has been proposed is e.g. lacunarity [6.] which is a measure of non-uniformity (or heterogeneity) of the structure [7.].
Sometimes the statistical-physics concepts like scaling and universality can be applied [8.] to identify the essential characteristics of a natural phenomenon. For example, as we will show in the preliminary applications, we will look for characteristic scale factors that can differ from cell to cell, but lead to a data collapse of the graphs corresponding to different cells in the population. If all the graphs after such rescaling can be collapsed onto basically one graph, this graph corresponds to a universal function and is an essential characteristics of the population under observation.
The result of tracing an individual neuronal structure with Neurolucida can be stored in DXF format where an individual dendritic branch is presented as a series of three--dimensional coordinates. The whole cell with the dendritic structure is presented in Fig. 1. In order to make the three-dimensional branches smoother and the discrete points along the branch equally spaced, we interpolate between the originally acquired points. At the end, each branch is characterized by a new set of equally spaced three-dimensional coordinates as shown in Fig. 2.
|
|
|
| Fig. 1. Original x-y-z coordinates of the dendritic tree of a typical dentate granule cell. The original points are connected by lines. | Fig. 2. After interpolation the x-y-z coordinates of each branch are equally spaced along the locus of the branch. |
We analyze the three-dimensional branching structure in the
following way: For each branch of the cell we determine its
branch order, its "parent" (i.e. the branch that it descends
from), its curvilinear length, L, and its end-to-end
distance, D. The ratio D/L is a measure of curvature of the
branch, and we will refer to this quantity hereafter as
straightness. By definition straightness D/L is
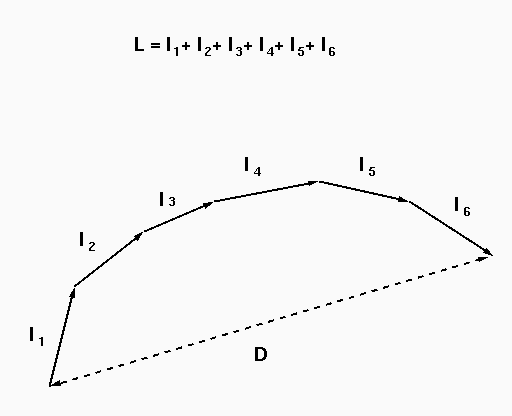
smaller or equal to 1. The more curved the branch is, the
smaller its straightness D/L as presented in Fig. A. For
each cell we then calculate an average branch length,
|
| Fig. A. Definitions of curvilinear length L and end-to-end distance D for a given branch. |
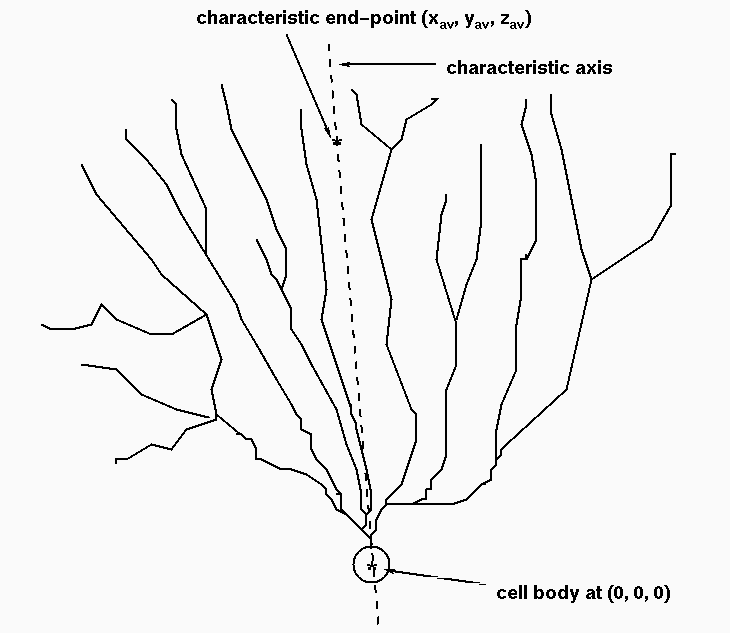
In addition, we analyze the dendritic end-points of a given dendritic structure. Particularly we focus on the spatial span of these end points. We determine the characteristic end-point (x_av, y_av, z_av) as an average over all end-points. The characteristic axis is defined as a line that goes through the center of the cell and the characteristic end point as presented in Fig. B. For each end-point we then calculate its distance r from the cell body and the cone angle Theta from the characteristic axis. We then average over the distances to obtain r_av and Theta_av. A rough estimate of the surface area that the branch end-points cover is calculated as 2 Pi r_av^2 [1 - cos(Theta_av)]. To do this estimate we assume that the branches span homogeneously in all directions which may be a too rough estimate, especially if the sample preparation is such that the tissue shrinks drastically in one direction.
|
| Fig. B. A hypothetical dendritic tree in 3D, cut with a characteristic axis. The characteristic axis is a line that goes through the center of the cell body and the point (x_av, y_av, z_av) which is an average of all branch end-points of the dendritic tree. The distance d_av between the cell body and the point (x_av, y_av, z_av) is called a characteristic distance. |
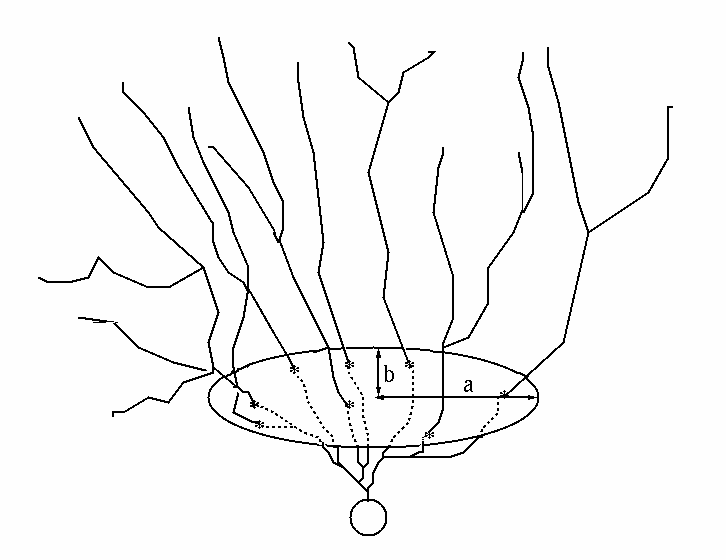
In order to be able to detect the differences between dendritic structures that are flat in one direction and spread in the other, we use the following 3D spatial anisotropy method. We cut the dendritic tree by parallel planes perpendicular to the characteristic axis. An individual plane is defined then by a distance d from the cell body. Within each plane we identify the points in which different branches intersect the plane. We determine the number N_INT of "chopped" branches as well as calculate a characteristic ellipsis with its long axis a and short axis b as shown in Fig. C. We take the area of the characteristic ellipsis to be an approximate area spanned by branches at that particular distance d from the cell center.
|
| Fig. C. A hypothetical dendritic tree in 3D, cut by a plane at a distance d from soma. The figure shows a characteristic ellipsis with its axes, a and b. The points in which the branches intersect this plane are denoted by *. |
Here we compare the proposed 3D spatial anisotropy method to the Sholl method which is standard method for characterizing the branching structure of various kinds of cells. We selected three dentate granule cells that yield very similar Sholl graphs as shown in Fig. 3(a). Each Sholl graph shows the number of intersections in dependence on the radial distance r from the center of soma. At a given radial distance r, the Sholl graph does not tell how these intersections are distributed on the sphere with radius r. Fig. 3(b) shows the number of intersections in dependecne on the planar distance d from the center of soma. The two graphs, Fig. 3(a) and 3(b), are almost identical. The reason for this is that the three dentate granule cells have branches that are on average more or less directed "upwards" (perpendicular to the pial surface???), such that cutting them with a plane is very similar to cutting them with a sphere.
While Sholl analysis gives sufficient information about the branching structure for cells that are spherically symmetric, it can not distinguish between spherically symmetric cells and cells with branches that are oriented preferentially along one particular direction, which is the case of dentate granule cells as well as CA1 pyramidal cells that we propose to study in the future. A more complete characterization of these non-spherically symmetric cells should include also quantities that characterize the distribution of branches around the center of cell body.
We therefore apply the 3D spatial anisotropy analysis to the three cells. Fig. 3(c) shows the ratio b/a of the short elliptic axis b versus the long elliptic axis a in dependence on the planar distance d from the cell body. First, we notice that the three graphs corresponding to the three different cells differ from one another drastically in contrast to Sholl graphs: The difference in b/a between the two extreme cells is more than a factor of 20. Second, even without comparison we can see that the ratio b/a is very small. Even in the best case the short axis b is more than 10 times smaller compared to the long axis a. This tells that the cells are not cylindrically symmetric around their characteristic axes. Fig. 3(d) shows the estimated cumulative volume covered by the dendritic tree, calculated by summing up all the elliptic areas: Volume = Sum {d1, d2,.... dN} Pi a b d_step. We can conlcude that the three cells cover very different volume

|
| Fig. 3. The comparison of methods on three different dentate granule cells that yield similar Sholl graphs. The Sholl graph (a) is compared to a similar analysis of number of intersections in dependence on the planar distance d from the cell (b). Both methods yield very similar results for all three cells. Graphs (c) and (d) are the results of 3D spatial anisotropy analysis: ratio b/a, of short axis b and long axis a of the characteristic ellipsis in dependence on the planar distance d from the cell body (c) and an estimated cumulative volume of the whole tree in dependence on the planar distance from the cell body (d). |
We average all these quantities over all aged and young adult animal cells separatelly. Our results show that the average branch length is 15% smaller in aged animals as compared to young adults. We find a similar result on examining end-to-end distances of branches. Further, we examine the average length of the branch in dependence on its branch order for each cell separately.
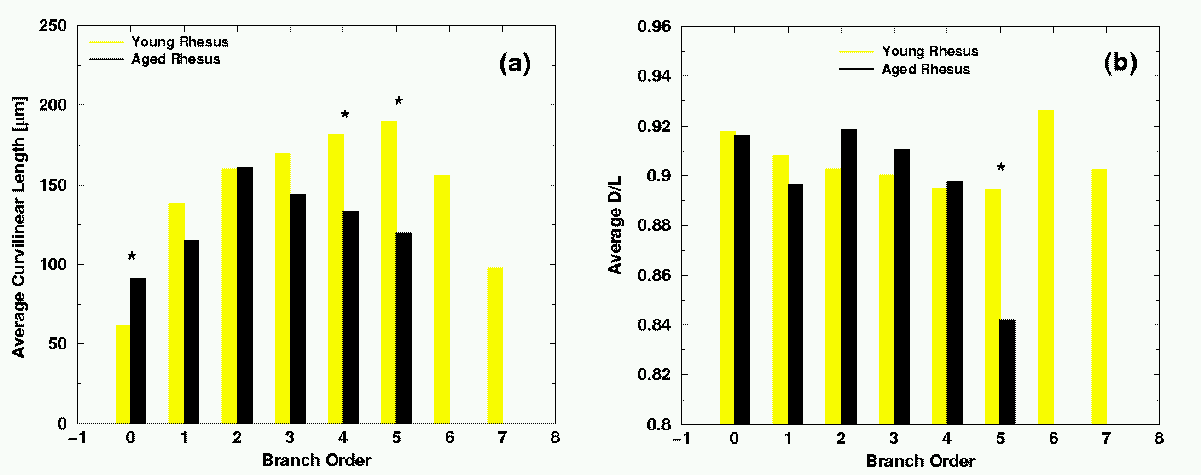
The results on Fig. 4(a) show that the average branch length of the proximal branches (branch orders 0, 1, 2, and 3) of aged animals is very close to the average length of young adult animals, while the average length of the distal branches (branch orders 4 and 5) is substantially smaller in aged rhesus monkey cells as compared to the cells of young adult animals.
We also study the ratio D/L, end-to-end distance (D) over curvilinear length (L), for each branch separately, which measures the degree of curvature of the branch. We average D/L for each branching order and plot the average D/L for young and aged cells, respectively. The results are presented in Fig. 4(b).
|
| Fig. 4. (a) Average length L of branches versus the branch order in dentate granule cells of young adult and aged rhesus monkeys. (b) The average ratio D/L of branches versus the branch order for young adult and aged rhesus monkeys. Bars with significant difference between the two groups are marked by *. |
In addition, we examine the end branch points in order to determine the spatial extent of the dendritic structure from the soma. Our results show that in young adult animals the end-branches reach 16% further as compared to aged animals as seen in Fig. 5.

|
| Fig. 5. Histograms of radia of the end-branch points that correspond to young adult and aged rhesus monkeys, respectively. We performed Kolmogorov-Smirnov test to find out that the two histograms differ from each other with a confidence level 92%. |
The above preliminary results show that the dendritic branches from aged animals are on average 15% shorter as compared to young adult animals. Proximal branches of young and aged animals do not seem to differ much in length, but the distal branches are significantly shorter in aged animals. As a result, the dendritic end branches reach 16% further away from the cell body in young adult as compared to aged animals.
We applied the 3D analysis of the number of branches versus the planar distance from the cell body in order to show how to distinguish the average morphology of young adult and aged cells. In addition, we will apply statistical physics concept of scaling to show how to extract the essential information from the data.
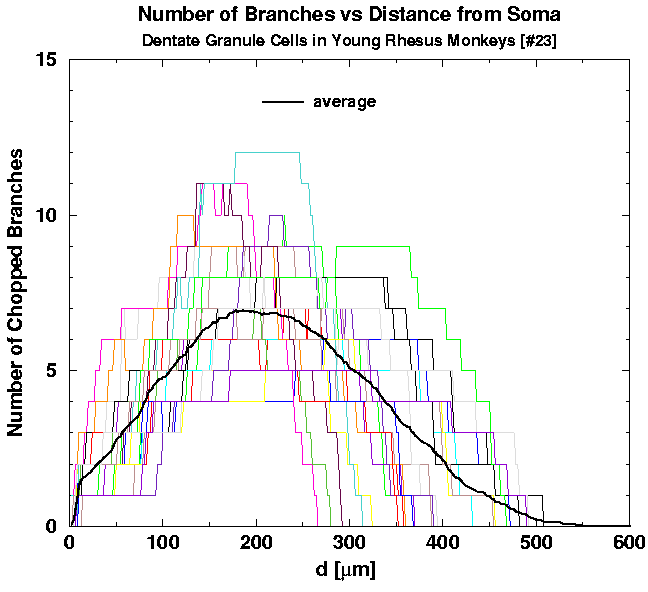
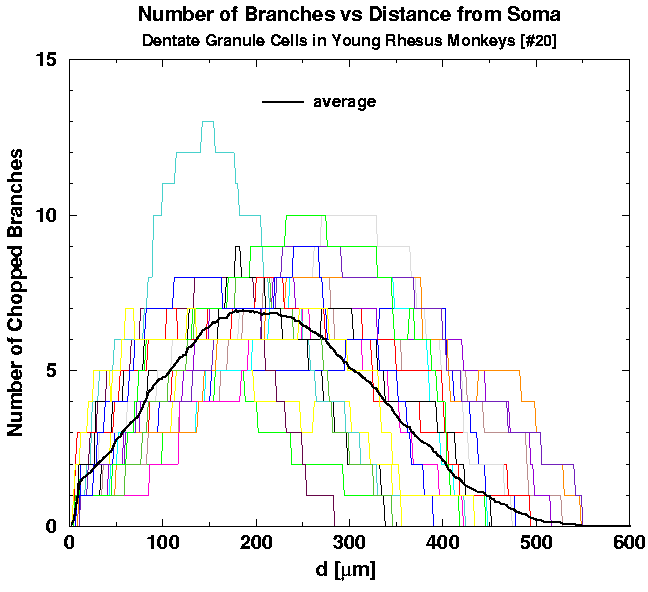
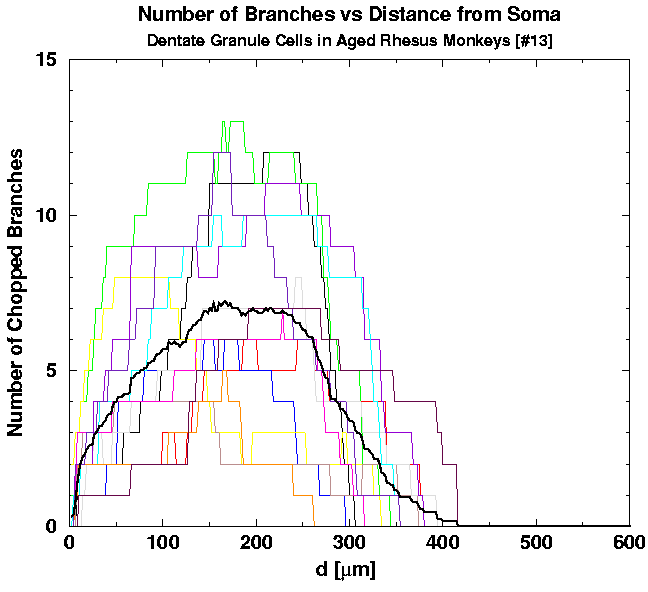
First, we plot the individual graphs of the number of intersections N_INT versus the planar distance d from the cell, for the two groups of young [Figs. 6(a) and 6(b)] and one group of aged cells [Fig. 6(c)]. On top of individual graphs we also plot the two averages, of all the young adult and all the aged cells (bold curves on the graphs). Although the two averages seem to be smooth functions, cell-to-cell variability is too huge to conclude either that the two averages differ significantly or not.
|
|
|
| Fig. 6a. Individual graphs of N_INT versus the distance d for 23 young cells. The average of all 43 young cells is shown for comparison. | Fig. 6b. Individual graphs of N_INT versus the distance d for 20 young cells. The average of all 43 young cells is shown for comparison. | Fig. 6c. Individual graphs of N_INT versus the distance d for 13 cells belonging to the aged animals. The average of all 13 graphs is shown for comparison. |
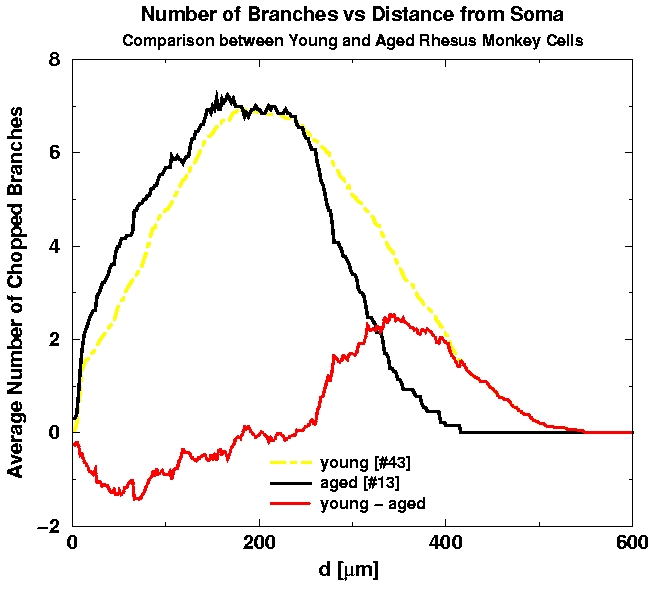
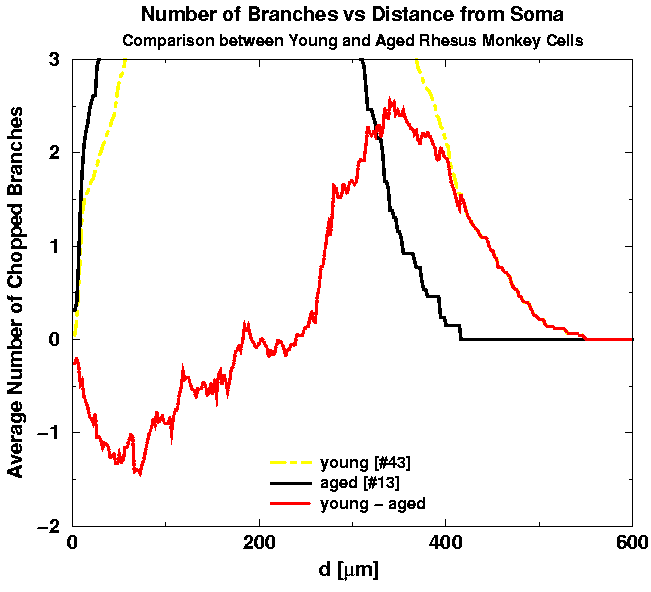
For clarity, the two average graphs are presented separately in Fig. 7(a) with their difference which is magnified in Fig. 7(b).
|
|
| Fig. 7a. Comparison of the two average graphs for young versus aged cells. | Fig. 7b. The difference between the two graphs, young and aged, is positive for distances d where there are more branches in young and negative for distances where there are more branches in aged cells. |
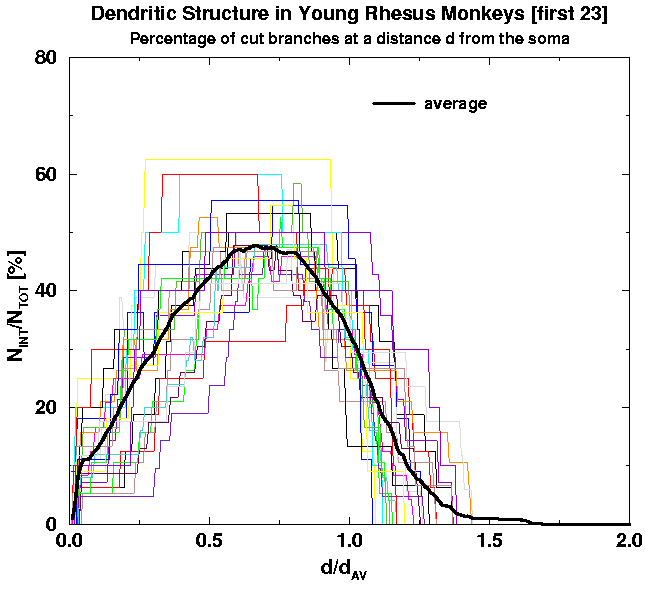
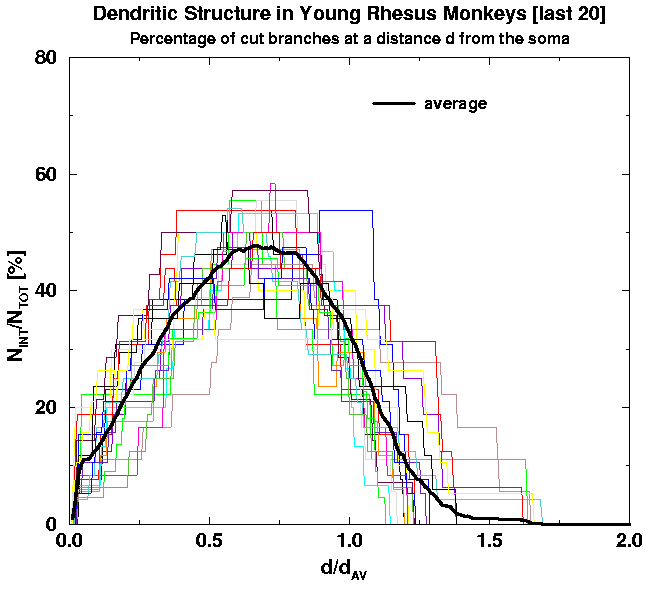
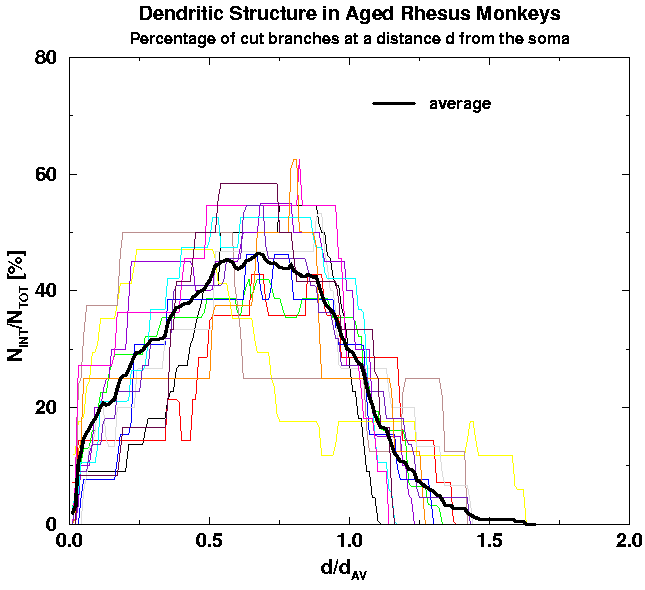
Second, we apply the scaling: We replot the above graphs in rescaled units in order to minimize cell-to-cell variability and with it the standard deviation of the averaged curves. We thus introduce the scaling factors such that individual curves corresponding to different cells would have the least cell-to-cell variability. The natural scaling factors are the characteristic distance d_av (which is a characteristic distance of the average of branch end-points from the cell body) and the total number of branches per cell, N_TOT, which is also a characteritic of an individual cell. In Fig. 8 we show the scaled versions of the graphs in Fig. 6. Comparing the graphs with the averages it is obvious to a naked eye that there is less cell-to-cell variability and thus the standard deviation of the averages gets correspondingly smaller.
|
|
|
| Fig. 8a. Rescalled Fig. 6a. Note much smaller fluctuations from cell to cell as compared to Fig. 6a. | Fig. 8b. Rescalled Fig. 6b. Note much smaller fluctuations from cell to cell as compared to Fig. 6b. | Fig. 8c. Rescalled Fig. 6c. Note much smaller fluctuations from cell to cell as compared to Fig. 6c. |
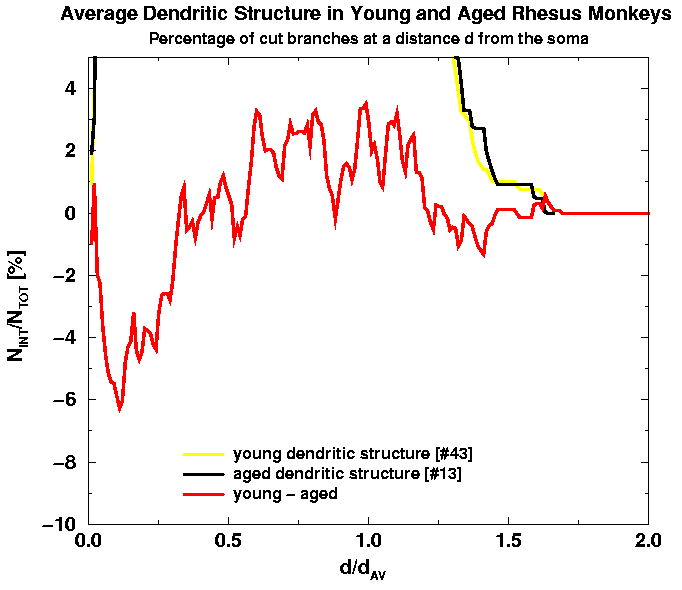
In Fig. 9(a) we again plot the two averaged graphs together with their difference which is magnified in Fig. 9(b).

|
|
| Fig. 9(a). Rescalled Fig. 7(a). Note much smaller fluctuations from cell to cell as compared to Fig. 7(a). | Fig. 9(b). Rescalled Fig. 7(b). Note much smaller fluctuations from cell to cell as compared to Fig. 7(b). |
We can conclude that the scaled form of the number of branch intersections, N_INT/N_TOT, versus the planar distance d/d_av has a well defined form that reminds a gaussian, truncated at d = 0, in the case of young adult cells, while it becomes asymmetric with relatively more branches at proximal distances, d < d_av/2, and relatively less branches at distal distances, d_av/2 < d 3d_av/2 in the case of aged cells. Whether these two averages from respective populations represent an universal function, characteristic of the population, still remains to be verified by using a better statistics, i.e. analyze around 100 cells per population. The above result is consistent with previous findings about the complexity of the branches in aged cells that seems to be higher as compared to young cells in the proximity of the cell body.
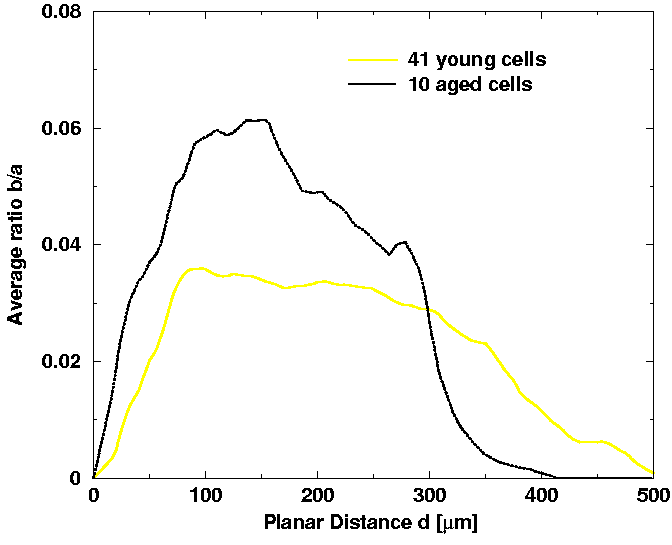
We also applied the 3D spatial anisotropy analysis to the dendritic structures of young adult and aged cells. Specifically, we analyzed the ratio of the two elliptic axis, b/a, in dependence on the planar distance d from the cell body. The results are presented in Fig. 10. In addition, we calculated the volume covered by the cell and we found no significant differences between the two population. Both results together lead to the following conclusions: The aged cells seem to have more spatially extended branches closer to the cell body as compared to young ones. However, the young ones have extended branches also in the distal part, in contrast to the aged cells. This is consistent with the other results found above: (a) higher branching complexity of aged cell at proximal distances as compared to young ones, and (b) dendritic structure of young cells extends approximately 16% further compared to dendritic structure of aged cells. Unfortunatelly, the differnces of the two graphs in Fig. 10 have still to be quantified. Due to a large cell-to-cell variations, we need to analyze in future of the order of 100 cells per population in order to quantify significant differences between the two populations.
|
| Fig. 10. The ratio of the two elliptic axes, b/a, in dependence on the planar distance d from the cell body, averaged over all the young adult cells and aged cells separately. |